Белок: строение и функции. Свойства белков
Белки — высокомолекулярные органические соединения, состоящие из остатков аминокислот, соединённых в длинную цепочку пептидной связью.
В состав белков живых организмов входит всего 20 типов аминокислот, все из которых относятся к альфа-аминокислотами, а аминокислотный состав белков и их порядок соединения друг с другом определяются индивидуальным генетическим кодом живого организма.
Одной из особенностей белков является их способность самопроизвольно формировать пространственные структуры характерные только для данного конкретного белка.
Из-за специфики своего строения белки могут обладать разнообразными свойствами. Например, белки, имеющие глобулярную четвертичную структуру, в частности белок куриного яйца, растворяются в воде с образованием коллоидных растворов. Белки, обладающие фибриллярной четвертичной структурой в воде не растворяются. Фибриллярными белками, в частности, образованы ногти, волосы, хрящи.
Химические свойства белков
Гидролиз
Все белки способны вступать в реакцию гидролиза. В случае полного гидролиза белков образуется смесь из α -аминокислот:
Белок + nH 2 O => смесь из α-аминокислот
Денатурация
Разрушение вторичной, третичной и четвертичной структур белка без разрушения его первичной структуры называют денатурацией. Денатурация белка может протекать под действием растворов солей натрия, калия или аммония – такая денатурация является обратимой:
Денатурация же протекающая под действием излучения (например, нагрева) или обработке белка солями тяжелых металлов является необратимой:
Так, например, необратимая денатурация белка наблюдается при термической обработке яиц в процессе их приготовления. В результате денатурации яичного белка его способность растворяться в воде с образованием коллоидного раствора исчезает.
Качественные реакции на белки
Биуретовая реакция
Если к раствору, содержащему белок добавить 10%-й раствор гидроксида натрия, а затем небольшое количество 1 %-го раствора сульфата меди, то появится фиолетовое окрашивание.
раствор белка + NаОН (10%-ный р-р) + СuSO 4 = фиолетовое окрашивание
Ксантопротеиновая реакция
растворы белка при кипячении с концентрированной азотной кислотой окрашиваются в желтый цвет:
раствор белка + HNO 3(конц.) => желтое окрашивание
Биологические функции белков
| каталитическая | ускоряют различные химические реакции в живых организмах | ферменты |
| структурная | строительный материал клеток | коллаген, белки клеточных мембран |
| защитная | защищают организм от инфекций | иммуноглобулины, интерферон |
| регуляторная | регулируют обменные процессы | гормоны |
| транспортная | перенос жизненно-необходимых веществ от одних частей организма к другим | гемоглобин переносит кислород |
| энергетическая | снабжают организм энергией | 1 грамм белка может обеспечить организм энергией в количестве 17,6 Дж |
| моторная (двигательная) | любые двигательные функции организма | миозин (мышечный белок) |
Среди органических веществ белки , или протеины , - самые многочисленные, наиболее разнообразные и имеющие первостепенное значение биополимеры. На их долю приходится 50 - 80% сухой массы клетки.
Молекулы белков имеют большие размеры, поэтому их называют макромолекулами . Кроме углерода , кислорода , водорода и азота , в состав белков могут входить сера, фосфор и железо. Белки отличаются друг от друга числом (от ста до нескольких тысяч), составом и последовательностью мономеров. Мономерами белков являются аминокислоты (рис. 1)
Бесконечное разнообразие белков создается за счет различного сочетания всего 20 аминокислот. Каждая аминокислота имеет свое название, особое строение и свойства. Их общую формулу можно представить в следующем виде:
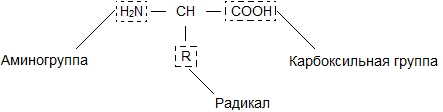
Молекула аминокислоты состоит из двух одинаковых для всех аминокислот частей, одна из которых является аминогруппой (-NH 2 ) с основными свойствами, другая - карбоксильной группой (-COOH ) с кислотными свойствами. Часть молекулы, называемая радикалом (R ), у разных аминокислот имеет различное строение. Наличие в одной молекуле аминокислоты основной и кислотной групп обусловливает их высокую реакционную способность. через эти группы происходит соединение аминокислот при образовании белка. При этом возникает молекула воды, а освободившиеся электроны образуют пептидную связь. Поэтому белки называют полипептидами .
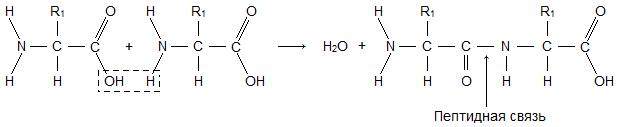
Молекулы белков могут иметь различные пространственные конфигурации, и в их строении различают четыре уровня структурной организации.
Последовательность аминокислот в составе полипептидной цепи представляет первичную структуру
белка. Она уникальна для любого белка и определяет его форму, свойства и функции.
Большинство белков имеют вид спирали в результате образования водородных связей между -CO-
и -NH-
группами разных аминокислотных остатков полипептидной цепи. Водородные связи малопрочные, но в комплексе они обеспечивают довольно прочную структуру. Эта спираль - вторичная структура
белка.
Третичная структура - трехмерная пространственная «упаковка» полипептидной цепи. В результате возникает причудливая, но для каждого белка специфическая конфигурация - глобула . Прочность третичной структуры обеспечивается разнообразными связями, возникающими между радикалами аминокислот.
Четвертичная структура
характерна не для всех белков. Она возникает в результате соединения нескольких макромолекул с третичной структурой в сложный комплекс. Например, гемоглобин крови человека представляет комплекс из четырех макромолекул белка.
Такая сложность структуры белковых молекул связана с разнообразием функций, свойственных этим биополимерам.
Нарушение природной структуры белка называют денатурацией
. Она может происходить под воздействием температуры, химических веществ, лучистой энергии и других факторов. При слабом воздействии распадается только четвертичная структура, при более сильном - третичная, а затем - вторичная, и белок остается в виде полипептидной цепи.
Этот процесс частично обратим: если не нарушена первичная структура, то денатурированный белок способен восстанавливать свою структуру. Отсюда следует, что все особенность строение макромолекулы белка определяются его первичной структурой.
Кроме простых белков , состоящих только из аминокислот, есть еще и сложные белки
БЕЛКИ (протеины), класс сложных азотсодержащих соединений, наиболее характерных и важных (наряду с нуклеиновыми кислотами) компонентов живого вещества. Белки выполняют многочисленные и разнообразные функции. Большинство белков ферменты, катализирующие химические реакции. Многие гормоны, регулирующие физиологические процессы, тоже являются белками. Такие структурные белки, как коллаген и кератин, служат главными компонентами костной ткани, волос и ногтей. Сократительные белки мышц обладают способностью изменять свою длину, используя химическую энергию для выполнения механической работы. К белкам относятся антитела, которые связывают и нейтрализуют токсичные вещества. Некоторые белки, способные реагировать на внешние воздействия (свет, запах), служат в органах чувств рецепторами, воспринимающими раздражение. Многие белки, расположенные внутри клетки и на клеточной мембране, выполняют регуляторные функции.В первой половине 19 в. многие химики, и среди них в первую очередь Ю.фон Либих, постепенно пришли к выводу, что белки представляют собой особый класс азотистых соединений. Название «протеины» (от греч.
protos первый) предложил в 1840 голландский химик Г.Мульдер. ФИЗИЧЕСКИЕ СВОЙСТВА Белки в твердом состоянии белого цвета, а в растворе бесцветны, если только они не несут какой-нибудь хромофорной (окрашенной) группы, как, например, гемоглобин. Растворимость в воде у разных белков сильно варьирует. Она изменяется также в зависимости от рН и от концентрации солей в растворе, так что можно подобрать условия, при которых один какой-нибудь белок будет избирательно осаждаться в присутствии других белков. Этот метод «высаливания» широко используется для выделения и очистки белков. Очищенный белок часто выпадает в осадок из раствора в виде кристаллов.В сравнении с другими соединениями молекулярная масса белков очень велика от нескольких тысяч до многих миллионов дальтон. Поэтому при ультрацентрифугировании белки осаждаются, и притом с разной скоростью. Благодаря присутствию в молекулах белков положительно и отрицательно заряженных групп они движутся с разной скоростью и в электрическом поле. На этом основан электрофорез метод, применяемый для выделения индивидуальных белков из сложных смесей. Очистку белков проводят и методом хроматографии.
ХИМИЧЕСКИЕ СВОЙСТВА Строение. Белки это полимеры, т.е. молекулы, построенные, как цепи, из повторяющихся мономерных звеньев, или субъединиц, роль которых играют у них a -аминокислоты. Общая формула аминокислот где R атом водорода или какая-нибудь органическая группа.Белковая молекула (полипептидная цепь) может состоять всего лишь из относительно небольшого числа аминокислот или из нескольких тысяч мономерных звеньев. Соединение аминокислот в цепи возможно потому, что у каждой из них имеются две разные химические группы: обладающая основными свойствами аминогруппа,
NH 2 , и кислотная карбоксильная группа, СООН. Обе эти группы присоединены к a -атому углерода. Карбоксильная группа одной аминокислоты может образовать амидную (пептидную) связь с аминогруппой другой аминокислоты: После того как две аминокислоты таким образом соединились, цепь может наращиваться путем добавления ко второй аминокислоте третьей и т.д. Как видно из приведенного выше уравнения, при образовании пептидной связи выделяется молекула воды. В присутствии кислот, щелочей или протеолитических ферментов реакция идет в обратном направлении: полипептидная цепь расщепляется на аминокислоты с присоединением воды. Такая реакция называется гидролизом. Гидролиз протекает спонтанно, а для соединения аминокислот в полипептидную цепь требуется энергия.
После того как две аминокислоты таким образом соединились, цепь может наращиваться путем добавления ко второй аминокислоте третьей и т.д. Как видно из приведенного выше уравнения, при образовании пептидной связи выделяется молекула воды. В присутствии кислот, щелочей или протеолитических ферментов реакция идет в обратном направлении: полипептидная цепь расщепляется на аминокислоты с присоединением воды. Такая реакция называется гидролизом. Гидролиз протекает спонтанно, а для соединения аминокислот в полипептидную цепь требуется энергия.
Карбоксильная группа и амидная группа (или сходная с ней имидная в случае аминокислоты пролина) имеются у всех аминокислот, различия же между аминокислотами определяются природой той группы, или «боковой цепи», которая обозначена выше буквой
R . Роль боковой цепи может играть и один атом водорода, как у аминокислоты глицина, и какая-нибудь объемистая группировка, как у гистидина и триптофана. Некоторые боковые цепи в химическом смысле инертны, тогда как другие обладают заметной реакционной способностью.Синтезировать можно многие тысячи различных аминокислот, и множество различных аминокислот встречается в природе, но для синтеза белков используется только 20 видов аминокислот: аланин, аргинин, аспарагин, аспарагиновая кислота, валин, гистидин, глицин, глутамин, глутаминовая кислота, изолейцин, лейцин, лизин, метионин, пролин, серин, тирозин, треонин, триптофан, фенилаланин и цистеин (в белках цистеин может присутствовать в виде димера
цистина). Правда, в некоторых белках присутствуют и другие аминокислоты, помимо регулярно встречающихся двадцати, но они образуются в результате модификации какой-нибудь из двадцати перечисленных уже после того, как она включилась в белок. Оптическая активность. У всех аминокислот, за исключением глицина, к a -атому углерода присоединены четыре разные группы. С точки зрения геометрии, четыре разные группы могут быть присоединены двумя способами, и соответственно есть две возможные конфигурации, или два изомера, относящиеся друг к другу, как предмет к своему зеркальному отражению, т.е. как левая рука к правой. Одну конфигурацию называют левой, или левовращающей ( L ), а другую правой, или правовращающей ( D ), поскольку два таких изомера различаются направлением вращения плоскости поляризованного света. В белках встречаются только L -аминокислоты (исключение составляет глицин; он может быть представлен лишь одной формой, поскольку у него две из четырех групп одинаковы), и все они обладают оптической активностью (поскольку имеется только один изомер). D -аминокислоты в природе редки; они встречаются в некоторых антибиотиках и клеточной оболочке бактерий. Последовательность аминокислот. Аминокислоты в полипептидной цепи располагаются не случайным образом, а в определенном фиксированном порядке, и именно этот порядок определяет функции и свойства белка. Варьируя порядок расположения 20 видов аминокислот, можно получить огромное число разных белков, точно так же, как из букв алфавита можно составить множество разных текстов.В прошлом на определение аминокислотной последовательности какого-нибудь белка уходило нередко несколько лет. Прямое определение и теперь достаточно трудоемкое дело, хотя созданы приборы, позволяющие вести его автоматически. Обычно проще бывает определить нуклеотидную последовательность соответствующего гена и вывести из нее аминокислотную последовательность белка. К настоящему времени уже определены аминокислотные последовательности многих сотен белков. Функции расшифрованных белков, как правило, известны, и это помогает представить себе возможные функции сходных белков, образующихся, например, при злокачественных новообразованиях.
Сложные белки. Белки, состоящие из одних только аминокислот, называют простыми. Часто, однако, к полипептидной цепи бывают присоединены атом металла или какое-нибудь химическое соединение, не являющееся аминокислотой. Такие белки называются сложными. Примером может служить гемоглобин: он содержит железопорфирин, который определяет его красный цвет и позволяет ему играть роль переносчика кислорода.В названиях большинства сложных белков содержится указание на природу присоединенных групп: в гликопротеинах присутствуют сахара, в липопротеинах жиры. Если от присоединенной группы зависит каталитическая активность фермента, то ее называют простетической группой. Нередко какой-нибудь витамин играет роль простетической группы или входит в ее состав. Витамин А, например, присоединенный к одному из белков сетчатки, определяет ее чувствительность к свету.
Третичная структура. Важна не столько сама аминокислотная последовательность белка (первичная структура), сколько способ ее укладки в пространстве. По всей длине полипептидной цепи ионы водорода образуют регулярные водородные связи, которые придают ей форму спирали либо слоя (вторичная структура). Из комбинации таких спиралей и слоев возникает компактная форма следующего порядка третичная структура белка. Вокруг связей, удерживающих мономерные звенья цепи, возможны повороты на небольшие углы. Поэтому с чисто геометрической точки зрения число возможных конфигураций для любой полипептидной цепи бесконечно велико. В действительности же каждый белок существует в норме только в одной конфигурации, определяемой его аминокислотной последовательностью. Структура эта не жесткая, она как бы « дышит» колеблется вокруг некой средней конфигурации. Цепь складывается в такую конфигурацию, при которой свободная энергия (способность производить работу) минимальна, подобно тому как отпущенная пружина сжимается лишь до состояния, соответствующего минимуму свободной энергии. Нередко одна часть цепи бывает жестко сцеплена с другой дисульфидными ( SS) связями между двумя остатками цистеина. Отчасти именно поэтому цистеин среди аминокислот играет особо важную роль.Сложность строения белков столь велика, что пока еще невозможно вычислить третичную структуру белка, если даже известна его аминокислотная последовательность. Но если удается получить кристаллы белка, то его третичную структуру можно определить по дифракции рентгеновских лучей.
У структурных, сократительных и некоторых других белков цепи вытянуты и несколько лежащих рядом слегка свернутых цепей образуют фибриллы; фибриллы, в свою очередь, складываются в более крупные образования волокна. Однако большинство белков в растворе имеет глобулярную форму: цепи свернуты в глобуле, как пряжа в клубке. Свободная энергия при такой конфигурации минимальна, поскольку гидрофобные («отталкивающие воду») аминокислоты скрыты внутри глобулы, а гидрофильные («притягивающие воду») находятся на ее поверхности.
Многие белки это комплексы из нескольких полипептидных цепей. Такое строение называется четвертичной структурой белка. Молекула гемоглобина, например, состоит из четырех субъединиц, каждая из которых представляет собой глобулярный белок.
Структурные белки благодаря своей линейной конфигурации образуют волокна, у которых предел прочности на разрыв очень высок, глобулярная же конфигурация позволяет белкам вступать в специфические взаимодействия с другими соединениями. На поверхности глобулы при правильной укладке цепей возникают определенной формы полости, в которых размещены реакционноспособные химические группы. Если данный белок фермент, то другая, обычно меньшая, молекула какого-то вещества входит в такую полость подобно тому, как ключ входит в замок; при этом меняется конфигурация электронного облака молекулы под влиянием находящихся в полости химических групп, и это вынуждает ее определенным образом реагировать. Таким способом фермент катализирует реакцию. В молекулах антител тоже имеются полости, в которых различные чужеродные вещества связываются и тем самым обезвреживаются. Модель «ключа и замка», объясняющая взаимодействие белков с другими соединениями, позволяет понять специфичность ферментов и антител, т.е. их способность реагировать только с определенными соединениями.
Белки у разных видов организмов. Белки, выполняющие одну и ту же функцию у разных видов растений и животных и потому носящие одно и то же название, имеют и сходную конфигурацию. Они, однако, несколько различаются по своей аминокислотной последовательности. По мере того как виды дивергируют от общего предка, некоторые аминокислоты в определенных положениях замещаются в результате мутаций другими. Вредные мутации, являющиеся причиной наследственных болезней, выбраковываются естественным отбором, но полезные или по крайней мере нейтральные могут сохраняться. Чем ближе друг к другу два каких-нибудь биологических вида, тем меньше различий обнаруживается в их белках.Некоторые белки меняются относительно быстро, другие весьма консервативны. К последним принадлежит, например, цитохром с дыхательный фермент, имеющийся у большинства живых организмов. У человека и шимпанзе его аминокислотные последовательности идентичны, а в цитохроме с пшеницы иными оказались лишь 38% аминокислот. Даже сравнивая человека и бактерии, сходство цитохромов с (различия затрагивают здесь 65% аминокислот) все еще можно заметить, хотя общий предок бактерии и человека жил на Земле около двух миллиардов лет назад. В наше время сравнение аминокислотных последовательностей часто используют для построения филогенетического (генеалогического) древа, отражающего эволюционные связи между разными организмами.
Денатурация. Синтезированная молекула белка, складываясь, приобретает свойственную ей конфигурацию. Эта конфигурация, однако, может разрушиться при нагревании, при изменении рН, под действием органических растворителей и даже при простом взбалтывании раствора до появления на его поверхности пузырьков. Измененный таким образом белок называют денатурированным; он утрачивает свою биологическую активность и обычно становится нерастворимым. Хорошо знакомые всем примеры денатурированного белка вареные яйца или взбитые сливки. Небольшие белки, содержащие всего лишь около сотни аминокислот, способны ренатурировать, т.е. вновь приобретать исходную конфигурацию. Но большинство белков превращается при этом просто в массу спутанных полипептидных цепей и прежнюю конфигурацию не восстанавливает.Одна из главных трудностей при выделении активных белков связана с их крайней чувствительностью к денатурации. Полезное применение это свойство белков находит при консервировании пищевых продуктов: высокая температура необратимо денатурирует ферменты микроорганизмов, и микроорганизмы погибают.
СИНТЕЗ БЕЛКОВ Для синтеза белка живой организм должен располагать системой ферментов, способных присоединять одну аминокислоту к другой. Необходим также источник информации, которая бы определяла, какие именно аминокислоты следует соединять. Поскольку в организме имеются тысячи видов белков и каждый из них состоит в среднем из нескольких сотен аминокислот, необходимая информация должна быть поистине огромной. Хранится она (подобно тому, как хранится запись на магнитной ленте) в молекулах нуклеиновых кислот, из которых состоят гены. См . также НАСЛЕДСТВЕННОСТЬ; НУКЛЕИНОВЫЕ КИСЛОТЫ. Активация ферментов. Синтезированная из аминокислот полипептидная цепь это далеко не всегда белок в его окончательной форме. Многие ферменты синтезируются сначала в виде неактивных предшественников и переходят в активную форму лишь после того, как другой фермент удалит на одном из концов цепи несколько аминокислот. В такой неактивной форме синтезируются некоторые из пищеварительных ферментов, например трипсин; эти ферменты активируются в пищеварительном тракте в результате удаления концевого фрагмента цепи. Гормон инсулин, молекула которого в активной форме состоит из двух коротких цепей, синтезируется в виде одной цепи, т.н. проинсулина. Затем средняя часть этой цепи удаляется, а оставшиеся фрагменты связываются друг с другом, образуя активную молекулу гормона. Сложные белки образуются лишь после того, как к белку будет присоединена определенная химическая группа, а для этого присоединения часто тоже требуется фермент. Метаболический кругооборот. После скармливания животному аминокислот, меченных радиоактивными изотопами углерода, азота или водорода, метка быстро включается в его белки. Если меченые аминокислоты перестают поступать в организм, то количество метки в белках начинает снижаться. Эти эксперименты показывают, что образовавшиеся белки не сохраняются в организме до конца жизни. Все они, за немногими исключениями, находятся в динамичном состоянии, постоянно распадаются до аминокислот, а затем вновь синтезируются.Некоторые белки распадаются, когда гибнут и разрушаются клетки. Это постоянно происходит, например, с эритроцитами и клетками эпителия, выстилающего внутреннюю поверхность кишечника. Кроме того, распад и ресинтез белков протекают и в живых клетках. Как ни странно, о распаде белков известно меньше, чем об их синтезе. Ясно, однако, что в распаде участвуют протеолитические ферменты, сходные с теми, которые расщепляют белки до аминокислот в пищеварительном тракте.
Период полураспада у разных белков различен от нескольких часов до многих месяцев. Единственное исключение молекулы коллагена. Однажды образовавшись, они остаются стабильными, не обновляются и не замещаются. Со временем, однако, меняются некоторые их свойства, в частности эластичность, а поскольку они не обновляются, следствием этого оказываются определенные возрастные изменения, например появление морщин на коже.
Синтетические белки. Химики давно уже научились полимеризовать аминокислоты, но аминокислоты соединяются при этом неупорядоченно, так что продукты такой полимеризации мало похожи на природные. Правда, имеется возможность соединять аминокислоты в заданном порядке, что позволяет получать некоторые биологически активные белки, в частности инсулин. Процесс достаточно сложен, и таким способом удается получать лишь те белки, в молекулах которых содержится около сотни аминокислот. Предпочтительнее вместо этого синтезировать или выделить нуклеотидную последовательность гена, соответствующую желаемой аминокислотной последовательности, а затем ввести этот ген в бактерию, которая и будет вырабатывать путем репликации большое количество нужного продукта. У этого метода, впрочем, тоже есть свои недостатки. См . также ГЕННАЯ ИНЖЕНЕРИЯ. БЕЛКИ И ПИТАНИЕ Когда белки в организме распадаются до аминокислот, эти аминокислоты могут быть снова использованы для синтеза белков. В то же время и сами аминокислоты подвержены распаду, так что они реутилизируются не полностью. Ясно также, что в период роста, при беременности и заживлении ран синтез белков должен превышать распад. Некоторые же белки организм непрерывно теряет; это белки волос, ногтей и поверхностного слоя кожи. Поэтому для синтеза белков каждый организм должен получать аминокислоты с пищей. Зеленые растения синтезируют из СО 2 , воды и аммиака или нитратов все 20 аминокислот, встречающихся в белках. Многие бактерии тоже способны синтезировать аминокислоты при наличии сахара (или какого-нибудь его эквивалента) и фиксированного азота, но и сахар, в конечном счете, поставляется зелеными растениями. У животных способность к синтезу аминокислот ограниченна; они получают аминокислоты, поедая зеленые растения или других животных. В пищеварительном тракте поглощенные белки расщепляются до аминокислот, последние всасываются, и уже из них строятся белки, характерные для данного организма. Ни один поглощенный белок не включается в структуры тела как таковой. Единственное исключение заключается в том, что у многих млекопитающих часть материнских антител может в интактном виде попасть через плаценту в кровоток плода, а через материнское молоко (особенно у жвачных) быть передано новорожденному сразу же после его появления на свет. Потребность в белках. Ясно, что для поддержания жизни организм должен получать с пищей некоторое количество белков. Однако размеры этой потребности зависят от ряда факторов. Организму необходима пища и как источник энергии (калорий), и как материал для построения его структур. На первом месте стоит потребность в энергии. Это значит, что, когда углеводов и жиров в рационе мало, пищевые белки используются не для синтеза собственных белков, а в качестве источника калорий. При длительном голодании даже собственные белки расходуются на удовлетворение энергетических нужд. Если же углеводов в рационе достаточно, то потребление белков может быть снижено. Азотистый баланс. В среднем ок. 16% всей массы белка составляет азот. Когда входившие в состав белков аминокислоты расщепляются, содержавшийся в них азот выводится из организма с мочой и (в меньшей мере) с калом в виде различных азотистых соединений. Удобно поэтому для оценки качества белкового питания использовать такой показатель, как азотистый баланс, т.е. разность (в граммах) между количеством азота, поступившего в организм, и количеством выведенного азота за сутки. При нормальном питании у взрослого эти количества равны. У растущего организма количество выведенного азота меньше количества поступившего, т.е. баланс положителен. При нехватке белков в рационе баланс отрицателен. Если калорий в рационе достаточно, но белки в нем полностью отсутствуют, организм сберегает белки. Белковый обмен при этом замедляется, и повторная утилизация аминокислот в синтезе белка идет с максимально возможной эффективностью. Однако потери неизбежны, и азотистые соединения все же выводятся с мочой и частично с калом. Количество азота, выведенного из организма за сутки при белковом голодании, может служить мерой суточной нехватки белка. Естественно предположить, что, введя в рацион количество белка, эквивалентное этому дефициту, можно восстановить азотистый баланс. Однако это не так. Получив такое количество белка, организм начинает использовать аминокислоты менее эффективно, так что для восстановления азотистого баланса требуется некоторое дополнительное количество белка.Если количество белка в рационе превышает необходимое для поддержания азотистого баланса, то вреда от этого, по-видимому, нет. Избыток аминокислот просто используется как источник энергии. В качестве особенно яркого примера можно сослаться на эскимосов, которые потребляют мало углеводов и примерно в десять раз больше белка, чем требуется для поддержания азотистого баланса. В большинстве случаев, однако, использование белка в качестве источника энергии невыгодно, поскольку из определенного количества углеводов можно получить намного больше калорий, чем из такого же количества белка. В бедных странах население получает необходимые калории за счет углеводов и потребляет минимальное количество белка.
Если необходимое число калорий организм получает в форме небелковых продуктов, то минимальное количество белка, обеспечивающее поддержание азотистого баланса, составляет для взрослого человека ок. 30 г в день. Примерно столько белка содержится в четырех ломтиках хлеба или 0,5 л молока. Оптимальным считают обычно несколько большее количество; рекомендуется от 50 до 70 г.
Незаменимые аминокислоты. До сих пор белок рассматривался как нечто целое. Между тем для того, чтобы мог идти синтез белка, в организме должны присутствовать все необходимые аминокислоты. Некоторые из аминокислот организм животного сам способен синтезировать. Их называют заменимыми, поскольку они не обязательно должны присутствовать в рационе, важно лишь, чтобы в целом поступление белка как источника азота было достаточным; тогда при нехватке заменимых аминокислот организм может синтезировать их за счет тех, что присутствуют в избытке. Остальные, «незаменимые», аминокислоты не могут быть синтезированы и должны поступать в организм с пищей. Для человека незаменимыми являются валин, лейцин, изолейцин, треонин, метионин, фенилаланин, триптофан, гистидин, лизин и аргинин. (Хотя аргинин и может синтезироваться в организме, его относят к незаменимым аминокислотам, поскольку у новорожденных и растущих детей он образуется в недостаточном количестве. С другой стороны, для человека зрелого возраста поступление некоторых из этих аминокислот с пищей может стать необязательным.)Этот список незаменимых аминокислот приблизительно одинаков также и у других позвоночных и даже у насекомых. Питательную ценность белков обычно определяют, скармливая их растущим крысам и следя за прибавкой веса животных.
Питательная ценность белков. Питательную ценность белка определяют по той незаменимой аминокислоте, которой более всего не хватает. Проиллюстрируем это на примере. В белках нашего тела содержится в среднем ок. 2% триптофана (по весу). Допустим, что в рацион входит 10 г белка, содержащего 1% триптофана, и что других незаменимых аминокислот в нем достаточно. В нашем случае 10 г этого неполноценного белка по сути эквивалентны 5 г полноценного; остальные 5 г могут послужить только источником энергии. Отметим, что, поскольку аминокислоты в организме практически не запасаются, а для того чтобы мог идти синтез белка, должны одновременно присутствовать все аминокислоты, эффект от поступления незаменимых аминокислот можно обнаружить лишь в том случае, если все они поступят в организм одновременно . Усредненный состав большей части животных белков близок к усредненному составу белков человеческого тела, так что аминокислотная недостаточность нам вряд ли грозит, если наш рацион богат такими продуктами, как мясо, яйца, молоко и сыр. Однако есть белки, например желатин (продукт денатурации коллагена), которые содержат очень мало незаменимых аминокислот. Растительные белки, хотя они в этом смысле и лучше желатина, тоже бедны незаменимыми аминокислотами; особенно мало в них лизина и триптофана. Тем не менее и чисто вегетарианскую диету вовсе нельзя считать вредной, если только при этом потребляется несколько большее количество растительных белков, достаточное для того, чтобы обеспечить организм незаменимыми аминокислотами. Больше всего белка содержится у растений в семенах, особенно в семенах пшеницы и различных бобовых культур. Богаты белком также и молодые побеги, например у спаржи. Синтетические белки в рационе. Добавляя небольшие количества синтетических незаменимых аминокислот или богатых ими белков к неполноценным белкам, например к белкам кукурузы, можно значительно повысить питательную ценность последних, т.е. тем самым как бы увеличить количество потребляемого белка. Другая возможность состоит в выращивании бактерий или дрожжей на углеводородах нефти с добавлением нитратов или аммиака в качестве источника азота. Полученный таким путем микробный белок может служить кормом для домашней птицы или скота, а может и непосредственно потребляться человеком. Третий, широко применяющийся, метод использует особенности физиологии жвачных животных. У жвачных в начальном отделе желудка, т.н. рубце, обитают особые формы бактерий и простейших, которые превращают неполноценные растительные белки в более полноценные микробные белки, а эти, в свою очередь, после переваривания и всасывания превращаются в животные белки. К корму скота можно добавить мочевину дешевое синтетическое азотсодержащее соединение. Обитающие в рубце микроорганизмы используют азот мочевины для превращения углеводов (которых в корме значительно больше) в белок. Около трети всего азота в корме скота может поступать в виде мочевины, что по сути и означает в определенной мере химический синтез белка. В США этот метод играет важную роль как один из способов получения белка. ЛИТЕРАТУРА Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека , тт. 12. М., 1993Албертс Б., Брей Д., Льюс Дж. и др. Молекулярная биология клетки , тт. 13. М., 1994
>> Химия: Белки
Белками, или белковыми веществами, называют высокомолекулярные (молекулярная масса варьирует от 5-10 тыс. до 1 млн и более) природные полимеры, молекулы которых построены, из остатков аминокислот, соединенных амидной (пептидной) связью.
Белки также называют протеинами (от греч. «протос» - первый, важный). Число остатков аминокислот в молекуле белка очень сильно колеблется и иногда достигает нескольких тысяч. Каждый белок обладает своей, присущей ему последовательностью расположения аминокислотных остатков.
Белки выполняют разнообразные биологические функции: каталитические (ферменты), регуляторные (гормоны), структурные (коллаген, фиброин), двигательные (миозин), транспортные (гемоглобин, миоглобин), защитные (иммуноглобулины, интерферон), запасные (казеин, альбумин, глиадин) и другие. Среди белков встречаются антибиотики и вещества, оказывающие токсическое действие.
Белки - основа биомембран, важнейшей составной части клетки и клеточных компонентов. Они играют ключевую роль в жизни клетки, составляя как бы материальную основу ее химической деятельности.
Исключительное свойство белка - самоорганизация структуры, т. е. его способность самопроизвольно создавать определенную, свойственную только данному белку пространственную структуру. По существу, вся деятельность организма (развитие, движение, выполнение им различных функций и многое другое) связана с белковыми веществами (рис. 36). Без белков невозможно представить себе жизнь.
Белки - важнейшая составная часть пищи человека и животных, поставщик необходимых им аминокислот
Строение
В пространственном строении белков большое значение имеет характер радикалов (остатков) R- в молекулах аминокислот. Неполярные радикалы аминокислот обычно располагаются внутри макромолекулы белка и обусловливают гидрофобные (см. ниже) взаимодействия; полярные радикалы, содержащие ионогенные (образующие ионы) группы, обычно находятся на поверхности макромолекулы белка и характеризуют электростатические (ионные) взаимодействия. Полярные не-ионогенные радикалы (например, содержащие спиртовые ОН-группы, амидные группы) могут располагаться как на поверхности, так и внутри белковой молекулы. Они участвуют в образовании водородных связей.
В молекулах белка а-аминокислоты связаны между собой пептидными (-СО-NH-) связями:
Построенные таким образом полипептидные цепи или отдельные участки внутри полипептидной цепи могут быть в отдельных случаях дополнительно связаны между собой дисуль-фидными (-S-S-) связями, или, как их часто называют, ди-сульфидными мостиками.
Большую роль в создании структуры белков играют ионные (солевые) и водородные связи, а также гидрофобное взаимодействие - особый вид контактов между гидрофобными компонентами молекул белков в водной среде. Все эти связи имеют различную прочность и обеспечивают образование сложной, большой молекулы белка.
Несмотря на различие в строении и функциях белковых веществ, их элементный состав колеблется незначительно (в % на сухую массу): углерода - 51-53; кислорода - 21,5- 23,5; азота - 16,8-18,4; водорода - 6,5-7,3; серы - 0,3-2,5. Некоторые белки содержат в небольших количествах фосфор, селен и другие элементы.
Последовательность соединения аминокислотных остатков в полипептидной цепи получила название первичной структурой белка (рис. 37).
Белковая молекула может состоять из одной или нескольких полипептидных цепей, каждая из которых содержит различное число аминокислотных остатков. Учитывая число их возможных комбинаций, можно сказать, что разнообразие белков почти безгранично, но не все из них существуют в природе. Общее число различных типов белков у всех видов живых организмов составляет 10 10 -10 12 . Для белков, строение которых отличается исключительной сложностью, кроме первичной, различают и более высокие уровни структурной организации: вторичную, третичную, а иногда и четвертичную структуры (табл. 9). Вторичной структурой обладает большая часть белков, правда, не всегда на всем протяжении полипептидной цепи. Полипептидные цепочки с определенной вторичной структурой могут быть по-разному расположены в пространстве.
Это пространственное расположение получило название третичной структуры (рис. 39)
В формировании третичной структуры, кроме водородных связей, большую роль играет ионное и гидрофобное взаимодействие. По характеру «упаковки» белковой молекулы различают глобулярные, или шаровидные, и фибриллярные, или нитевидные, белки.
Для глобулярных белков более характерна а-спиральная структура, спирали изогнуты, «свернуты». Макромолекула имеет сферическую форму. Они растворяются в воде и солевых растворах с образованием коллоидных систем. Большинство белков животных, растений и микроорганизмов относится к глобулярным белкам.
Для фибриллярных белков более характерна нитевидная структура. Они, как правило, не растворяются в воде. Фибриллярные белки обычно выполняют структурообразующие функции. Их свойства (прочность, способность растягиваться) зависят от способа упаковки полипептидных цепочек. Примером фибриллярных белков служат белки мускульной ткани (миозин), кератин (роговая ткань). В ряде случаев отдельные субъединицы белка с помощью водородных связей, электростатического и других взаимодействий образуют сложные ансамбли. В этом случае образуется четвертичная структура белков.
Однако следует еще раз отметить, что в организации более высоких структур белка исключительная роль принадлежит первичной структуре.
Классификация
Существует несколько классификаций белков. В их основе лежат разные признаки:
Степень сложности (простые и сложные);
Форма молекул (глобулярные и фибриллярные белки);
Растворимость в отдельных растворителях (водорастворимые, растворимые в разбавленных солевых растворах - альбумины, спирторастворимые - проламины, растворимые в разбавленных щелочах и кислотах - глутелины);
Выполняемая функция (например, запасные белки, скелетные и т. п.).
Свойства
Белки - амфотерные электролиты. При определенном значении рН среды (она называется изоэлектрической точкой) число положительных и отрицательных зарядов в молекуле белка одинаково. Это одно из основных свойств белка. Белки в этой точке электронейтральны, а их растворимость в воде наименьшая. Способность белков снижать растворимость при достижении электронейтральности их молекул используется для выделения их из растворов, например, в технологии получения белковых продуктов.
Гидратация
Процесс гидратации означает связывание белками воды, при этом они проявляют гидрофильные свойства: набухают, их масса и объем увеличиваются. Набухание белка сопровождается его частичным растворением. Гидрофильность отдельных белков зависит от их строения. Имеющиеся в составе и расположенные на поверхности белковой макромолекулы гидрофильные амидные (-СО-NН-, пептидная связь), аминные (NH2) и карбоксильные (СООН) группы притягивают к себе молекулы воды, строго ориентируя их на поверхности молекулы. Окружающая белковые глобулы гидратная (водная) оболочка препятствует агрегации и осаждению, а следовательно, способствует устойчивости растворов белка. В изоэлектрической точке белки обладают наименьшей способностью связывать воду, происходит разрушение гидратной оболочки вокруг белковых молекул, поэтому они соединяются, образуя крупные агрегаты. Агрегация белковых молекул происходит и при их обезвоживании с помощью некоторых органических растворителей, например этилового спирта. Это приводит к выпадению белков в осадок. При изменении рН среды макромолекула белка становится заряженной, и его гидратационная способность меняется.
При ограниченном набухании концентрированные белковые растворы образуют сложные системы, называемые студнями. Студни не текучи, упруги, обладают пластичностью, определенной механической прочностью, способны сохранять свою форму. Глобулярные белки могут полностью гидратироваться, растворяясь в воде (например, белки молока), образуя растворы с невысокой концентрацией. Гидрофильные свойства белков, т. е. их способность набухать, образовывать студни, стабилизировать суспензии, эмульсии и пены, имеют большое значение в биологии и пищевой промышленности . Очень подвижным студнем, построенным в основном из молекул белка, является цитоплазма - полужидкое содержимое клетки. Сильно гидратированный студень - сырая клейковина, выделенная из пшеничного теста, она содержит до 65% воды. Различная гидрофильность клейковинных белков - один из признаков, характеризующих качество зерна пшеницы и получаемой из него муки (так называемые сильные и слабые пшеницы). Гидрофильность белков зерна и муки играет большую роль при хранении и переработке зерна, в хлебопечении. Тесто, которое получают в хлебопекарном производстве, представляет собой набухший в воде белок, концентрированный студень, содержащий зерна крахмала.
Денатурация белков
При денатурации под влиянием внешних факторов (температуры, механического воздействия, действия химических агентов и ряда других факторов) происходит изменение вторичной, третичной и четвертичной структур белковой макромолекулы, т. е. ее нативной пространственной структуры. Первичная структура, а следовательно, и химический состав белка не меняются. Изменяются физические свойства: снижается растворимость, способность к гидратации, теряется биологическая активность. Меняется форма белковой макромолекулы, происходит агрегирование. В то же время увеличивается активность некоторых химических групп, облегчается воздействие на белки протеолитических ферментов, а следовательно, он легче гидролизуется.
В пищевой технологии особое практическое значение имеет тепловая денатурация белков, степень которой зависит от температуры, продолжительности нагрева и влажности. Это необходимо помнить при разработке режимов термообработки пищевого сырья, полуфабрикатов, а иногда и готовых продуктов. Особую роль процессы тепловой денатурации играют при бланшировании растительного сырья, сушке зерна, выпечке хлеба, получении макаронных изделий. Денатурация белков может вызываться и механическим воздействием (давлением, растиранием, встряхиванием, ультразвуком). Наконец, к денатурации белков приводит действие химических реагентов (кислот, щелочей, спирта, ацетона). Все эти приемы широко используют в пищевой и биотехнологии.
Пенообразов ание
Под процессом пенообразования понимают способность белков образовывать высококонцентрированные системы «жидкость - газ», называемые пенами. Устойчивость пены, в которой белок является пенообразователем, зависит не только от его природы и от концентрации, но и от температуры. Белки в качестве пенообразователей широко используются в кондитерской промышленности (пастила, зефир, суфле). Структуру пены имеет хлеб, а это влияет на его вкусовые свойства.
Молекулы белков под влиянием ряда факторов могут разрушаться или вступать во взаимодействие с другими веществами с образованием новых продуктов. Для пищевой промышленности можно выделить два очень важных процесса: 1) гидролиз белков под действием ферментов и 2) взаимодействие аминогрупп белков или аминокислот с карбонильными группами восстанавливающих Сахаров. Под влиянием протеаз - ферментов, катализирующих гидролитическое расщепление белков, последние распадаются на более простые продукты (поли- и дипептиды) и в конечном итоге на аминокислоты . Скорость гидролиза белка зависит от его состава, молекулярной структуры, активности фермента и условий.
Гидролиз белков
Реакцию гидролиза с образованием аминокислот в общем виде можно записать так:
Горение
4. С помощью каких реакций можно распознать белки?
5. Какую роль играют белки в жизни организмов?
6. Вспомните из курса общей биологии, какие белки определяют иммунные свойства организмов.
7. Расскажите о СПИДе и профилактике этого страшного заболевания.
8. Как распознать изделие из натуральной шерсти и искусственного волокна?
9. Напишите уравнение реакции гидролиза белков, имеющих общую формулу (-NН-СН-СО-)n.
l
R
Какое значение имеет этот процесс в биологии и как он используется в промышленности?
10. Напишите уравнения реакций, с помощью которых можно осуществить следующие переходы: этан -> этиловый спирт -> уксусный альдегид -> уксусная кислота -> хлоруксусная кислота -> аминоуксусная кислота -> полипептид.
кейсы по химии , задачи и решения , конспекты уроков
№1. Белки: пептидная связь, их обнаружение.
Белки – макромолекулы линейных полиамидов, образованных а-аминокислотами в результате реакции поликонденсации в биологических объектах.
Белки – это высокомолекулярные соединения, построенные из аминокислот . В создание белков участвует 20 аминокислот. Они связываются между собой в длинные цепи, которые образуют основу белковой молекулы большой молекулярной массы.
Функции белков в организме
Сочетание своеобразных химических и физических свойств белков обеспечивает именно этому классу органических соединений центральную роль в явлениях жизни.
Белки имеют следующие биологические свойства, или осуществляют следующие основные функции в живых организмах:
1. Каталитическая функция белков. Все биологические катализаторы - ферменты являются белками. В настоящее время охарактеризовано тысячи ферментов, многие из них выделены в кристаллической форме. Почти все ферменты - мощные катализаторы, повышающие скорости реакций, по крайней мере, в миллион раз. Эта функция белков является уникальной, не свойственной другим полимерным молекулам.
2. Питательная (резервная функция белков). Это, прежде всего белки, предназначенные для питания развивающегося зародыша: казеин молока, овальбумин яиц, запасные белки семян растений. Ряд других белков, несомненно, используется в организме в качестве источника аминокислот, которые, в свою очередь, являются предшественниками биологически активных веществ, регулирующих процесс обмена веществ.
3. Транспортная функция белков. Транспорт многих небольших молекул и ионов осуществляется специфическими белками. Например, дыхательная функция крови, а именно перенос кислорода, выполняется молекулами гемоглобина - белка эритроцитов. В транспорте липидов принимают участие альбумины сыворотки крови. Ряд других сывороточных белков образует комплексы с жирами, медью, железом, тироксином, витамином А и другими соединениями, обеспечивая их доставку в соответствующие органы.
4. Защитная функция белков. Основную функцию защиты выполняет иммуннологическая система, которая обеспечивает синтез специфических защитных белков - антител - в ответ на поступление в организм бактерий, токсинов или вирусов (антигенов). Антитела связывают антигены, взаимодействуя с ними, и тем самым нейтрализуют их биологическое действие и сохраняют нормальное состояние организма. Свертывание белка плазмы крови - фибриногена - и образование сгустка крови, предохраняющего от потери крови при ранениях - еще один пример защитной функции белков.
5. Сократительная функция белков. В акте мышечного сокращения и расслабления участвует множество белков. Главную роль в этих процессах играют актин и миозин - специфические белки мышечной ткани. Сократительная функция присуща также и белкам субклеточных структур, что обеспечивает тончайшие процессы жизнедеятельности клеток,
6. Структурная функция белков. Белки с такой функцией занимают первое место среди других белков тела человека. Широко распространены такие структурные белки, как коллаген в соединительной ткани; кератин в волосах, ногтях, коже; эластин - в сосудистых стенках и др.
7. Гормональная (регуляторная) функция белков. Обмен веществ в организме регулируется разнообразными механизмами. В этой регуляцииважное место занимают гормоны, вырабатываемые железами внутреннейсекреции. Ряд гормонов представлен белками, или полипептидами, например гормоны гипофиза, поджелудочной железы и др.
Пептидная связь
Формально образование белковой макромолекулы можно представить как реакцию поликонденсации α-аминокислот.
С химической точки зрения белки - это высокомолекулярные азотсодержащие органические соединения (полиамиды), молекулы которых построены из остатков аминокислот. Мономерами белков служат α-аминокислоты, общим признаком которых является наличие карбоксильной группы -СООН и аминогруппы -NH 2 у второго углеродного атома (α-углеродный атом):
Исходя из результатов изучения продуктов гидролиза белков и выдвинутых А.Я. Данилевским идей о роли пептидных связей -CO-NH- в построении белковой молекулы, немецкий ученый Э.Фишер предложил в начале XX века пептидную теорию строения белков. Согласно этой теории, белки представляют собой линейные полимеры α-аминокислот, связанных пептидной связью - полипептиды:

В каждом пептиде один концевой аминокислотный остаток имеет свободную α-аминогруппу (N-конец), а другой - свободную α-карбоксильную группу (С-конец). Структуру пептидов принято изображать, начиная с N-концевой аминокислоты. При этом аминокислотные остатки обозначаются символами. Например: Ala-Tyr-Leu-Ser-Tyr- - Cys. Этой записью обозначен пептид, в котором N-концевой α-аминокислотой яв ляется аланин, а С-концевой - цистеин. При чтении такой записи окончания названий всех кислот, кроме последних меняются на - "ил": аланил-тирозил-лейцил-серил-тирозил- -цистеин. Длина пептидной цепи в пептидах и белках, встречающихся в организме, колеблется от двух до сотен и тысяч аминокислотных остатков.
№2. Классификация простых белков.
К простым (протеинам) относят белки, дающие при гидролизе только аминокислоты.
Протеиноиды ____простые белки животного происхождения, нерастворимые вводе, растворах солей, разбавленных кислотах и щелочах. Выполняют главным образом опорные функции (например, Коллаген, кератин
протамины – положительно заряженные ядерные белки, с молекулярной массой 10-12 kDa. Примерно на 80% состоят из щелочных аминокислот, что дает им возможность взаимодействовать с нуклеиновыми кислотами посредством ионных связей. Принимают участие в регуляции генной активности. Хорошо растворимы в воде;
гистоны – ядерные белки, играющие важную роль в регуляции генной активности. Они найдены во всех эукариотических клетках, и разделены на 5 классов, различающихся по молекулярной массе и аминокислотному. Молекулярная масса гистонов находится в интервале от 11 до 22 kDa, а различия в аминокислотном составе касаются лизина и аргинина, содержание которых варьирует от 11 до 29% и от 2 до 14% соответственно;
проламины – не растворимы в воде, но растворимы в 70% спирте, особенности хим.строения – много пролина, глутаминовой кислоты нет лизина,
глутелины – растворимы в щелочных растворах,
глобулины – белки, не растворимые в воде и в полунасыщенном растворе сернокислого аммония, но растворимые в водных растворах солей, щелочей и кислот. Молекулярная масса – 90-100 kDa;
альбумины – белки животных и растительных тканей, растворим в воде и солевых растворах. Молекулярнаяя масса равна 69 kDa;
склеропротеины – белки опорных тканей животных
В качестве примеров простых белков могут служить фиброин шелка, яичный сывороточный альбумин, пепсин и др.
№3. Способы выделения и осаждения (очистки) белков.





№4. Белки как полиэлектролиты. Изоэлектрическая точка белка.
Белки являются амфотерными полиэлектролитами, т.е. проявляют как кислотные, так и основные свойства. Это обусловлено наличием в молекулах белков аминокислотных радикалов, способных к ионизации, а также свободных α-амино- и α-карбоксильных групп на концах пептидных цепей. Кислотные свойства белку придают кислые аминокислоты (аспарагиновая, глутаминовая), а щелочные свойства - основные аминокислоты (лизин, аргинин, гистидин).
Заряд белковой молекулы зависит от ионизации кислых и основных групп аминокислотных радикалов. В зависимости от соотношения отрицательных и положительных групп молекула белка в целом приобретает суммарный положительный или отрицательный заряд. При подкислении раствора белка степень ионизации анионных групп снижается, а катионных повышается; при подщелачивании - наоборот. При определенном значении рН число положительно и отрицательно заряженных групп становится одинаковым, возникает изоэлектрическое состояние белка (суммарный заряд равен 0). Значение рН, при котором белок находится в изоэлектрическом состоянии, называют изоэлектрической точкой и обозначают pI, аналогично аминокислотам. Для большинства белков pI лежит в пределах 5,5-7,0, что свидетельствует о некотором преобладании в белках кислых аминокислот. Однако есть и щелочные белки, например, сальмин - основной белок из молок семги (pl=12). Кроме того, есть белки, у которых pI имеет очень низкое значение, например, пепсин - фермент желудочного сока (pl=l). В изоэлектрической точке белки очень неустойчивые и легко выпадают в осадок, обладая наименьшей растворимостью.
Если белок не находится в изоэлектрическом состоянии, то в электрическом поле его молекулы будут перемещаться к катоду или аноду, в зависимости от знака суммарного заряда и со скоростью, пропорциональной его величине; в этом заключается сущность метода электрофореза. Этим методом можно разделять белки с различным значением pI.
Белки хотя и обладают свойствами буфера, но емкость их при физиологических значениях рН ограничена. Исключение составляют белки, содержащие много гистидина, так как только радикал гистидина обладает буферными свойствами в интервале рН 6-8. Таких белков очень мало. Например, гемоглобин, содержащий почти 8% гистидина, является мощным внутриклеточным буфером в эритроцитах, поддерживая рН крови на постоянном уровне.
№5. Физико-химические свойства белков.
Белки имеют различные химические, физические и биологические свойства, которые определяются аминокислотным составом и пространственной организацией каждого белка. Химические реакции белков очень разнообразны, они обусловлены наличием NH 2 -, СООН-групп и радикалов различной природы. Это реакции нитрования, ацилирования, алкилирования, этерификации, окисления-восстановления и другие. Белки обладают кислотно-основными, буферными, коллоидными и осмотическими свойствами.
Кислотно-основные свойства белков
Химические свойства. При слабом нагревании водных растворов белков происходит денатурация. При этом образуется осадок.
При нагревании белков с кислотами происходит гидролиз, при этом образуется смесь аминокислот.
Физико-химические свойства белков
Белки имеют высокий молекулярный вес.
Заряд белковой молекулы. Все белки имеют хоть одну свободную -NH и - СООН группы.
Белковые растворы - коллоидные растворы с разными свойствами. Белки бывают кислыми и основными. Кислые белки содержат много глу и асп, у которых есть дополнительные карбоксильные и меньше аминогрупп. В щелочных белках много лиз и арг. Каждая молекула белка в водном растворе окружена гидратной оболочкой, так как у белков за счет аминокислот есть много гидрофильных группировок (-СООН, -ОН, -NH 2 , -SH). В водных растворах белковая молекула имеет заряд. Заряд белка в воде может меняться в зависимости от РН.
Осаждение белков. У белков есть гидратная оболочка, заряд, препятствующий склеиванию. Для осаждения необходимо снять гидратную оболочку и заряд.
1.Гидратация. Процесс гидратации означает связывание белками воды, при этом они проявляют гидрофильные свойства: набухают, их масса и объем увеличивается. Набухание белка сопровождается его частичным растворением. Гидрофильность отдельных белков зависит от их строения. Имеющиеся в составе и расположенные на поверхности белковой макромолекулы гидрофильные амидные (–CO–NH–, пептидная связь), аминные (NH2) и карбоксильные (COOH) группы притягивают к себе молекулы воды, строго ориентируя их на поверхность молекулы. Окружая белковые глобулы гидратная (водная) оболочка препятствует устойчивости растворов белка. В изоэлектрической точке белки обладают наименьшей способностью связывать воду, происходит разрушение гидратной оболочки вокруг белковых молекул, поэтому они соединяются, образуя крупные агрегаты. Агрегация белковых молекул происходит и при их обезвоживании с помощью некоторых органических растворителей, например этило- вого спирта. Это приводит к выпадению белков в осадок. При изменении pH среды макромолекула белка становится заряженной, и его гидратационная способность меняется.
Реакции осаждения делят на два вида.
Высаливание белков: (NH 4)SO 4 - снимается только гидратная оболочка, белок сохраняет все виды своей структуры, все связи, сохраняет нативные свойства. Такие белки можно затем вновь растворить и использовать.
Осаждения с потерей нативных свойств белка - процесс необратимый. С белка снимается гидратная оболочка и заряд, нарушаются различные свойства в белке. Например соли меди, ртути, мышьяка, железа, концентрированные неорганические кислоты - HNO 3 , H 2 SO 4 , HCl, органические кислоты, алкалоиды - танины, йодистая ртуть. Добавление органических растворителей понижает степень гидратации и приводит к осаждению белка. В качестве таких растворителей используют ацетон. Осаждают белки также с помощью солей, например, сульфата аммония. Принцип этого метода основан на том, что при повышении концентрации соли в растворе происходит сжатие ионных атмосфер, образуемых противоионами белка, что способствует сближению их до критического расстояния, на котором межмолекулярные силы ван-дер-ваальсова притяжения перевешивают кулоновские силы отталкивания противоионов. Это приводит к слипанию белковых частиц и их выпадению в осадок.
При кипячении молекулы белков начинают хаотично двигаться, сталкиваются, снимается заряд, уменьшается гидратная оболочка.
Для обнаружения белков в растворе применяются:
цветные реакции;
реакции осаждения.
Методы выделения и очистки белков.
высаливание;
электрофорез;
хроматография: адсорбция, расщепление;
ультрацентрифугирование.
гомогенизация - клетки растираются до однородной массы;
экстракция белков водными или водно-солевыми растворами;
Структурная организация белков.
Первичная структура - определяется последовательностью аминокислот в пептидной цепочке, стабилизируется ковалентными пептидными связями (инсулин, пепсин, химотрипсин).
Вторичная структура - пространственная структура белка. Это либо -спираль, либо -складчатость. Создаются водородные связи.
Третичная структура - глобулярные и фибриллярные белки. Стабилизируют водородные связи, электростатические силы (СОО-, NН3+), гидрофобные силы, сульфидные мостики, определяются первичной структурой. Глобулярные белки - все ферменты, гемоглобин, миоглобин. Фибриллярные белки - коллаген, миозин, актин.
Четвертичная структура - имеется только у некоторых белков. Такие белки построены из нескольких пептидов. Каждый пептид имеет свою первичную, вторичную, третичную структуру, называются протомерами. Несколько протомеров соединяются вместе в одну молекулу. Один протомер не функционирует как белок, а только в соединении с другими протомерами.
Пример: гемоглобин = -глобула + -глобула - переносит О 2 в совокупности, а не по раздельности.
Белок может ренатурировать. Для этого необходимо очень короткое воздействие агентов.

6) Способы обнаружения белков.
Белки – высокомолекулярные биологические полимеры, структурными (мономерными) звеньями которых служат -аминокислоты. Аминокислоты в белках соединены друг с другом пептидной связью,образование которой происходит за счет карбоксильной группы, стоящей у -углеродного атома одной аминокислоты и -аминной группы другой аминокислоты с выделением молекулы воды. Мономерные звенья белков называют остатками аминокислот.
Пептиды, полипептиды и белки отличаются не только количеством, составом но и последовательностью аминокислотных остатков, физико-химическими свойствами и функциями, выполняемыми в организме. Молекулярная масса белков варьирует от 6 тыс. до 1 млн. и более. Химические и физические свойства белков обусловлены химической природой и физико-химическими свойствами радикалов, входящих в них остатков аминокислот. Способы обнаружения и количественного определения белков в биологических объектах и продуктах питания, а также выделения их из тканей и биологических жидкостей основаны на физических и химических свойствах этих соединений.
Белки при взаимодействии с некоторыми химическими веществами дают окрашенные соединения . Образование этих соединений происходит при участии радикалов аминокислот, их специфических групп или пептидных связей. Цветные реакции позволяют установитьналичие белка в биологическом объекте или растворе и доказать присутствиеопределенных аминокислот в белковой молекуле . На основе цветных реакций разработаны некоторые методы количественного определения белков и аминокислот.
Универсальными считают биуретовую и нингидриновую реакции , так как их дают все белки.Ксантопротеиновая реакция, реакция Фоля и др. являются специфическими, так как они обусловлены радикальными группами определенных аминокислот в молекуле белка.
Цветные реакции позволяют установить наличие белка в исследуемом материале и присутствие определенных аминокислот в его молекулах.
Биуретовая реакция . Реакция обусловлена наличием в белках, пептидах, полипептидахпептидных связей , которые в щелочной среде образуют сионами меди (II) комплексные соединения, окрашенные вфиолетовый (с красным или с синим оттенком) цвет . Окраска обусловлена наличием в молекуле не менее двух групп-CO-NH- , связанных непосредственно между собой или при участии атома углерода или азота.
Ионы меди (II) соединяются двумя ионными связями с группами =С─О ˉ и четырьмя координационными связями с атомами азота (=N―).
Итенсивность окраски зависит от количества белка в растворе. Это позволяет использовать данную реакцию для количественного определения белка. Цвет окрашенных растворов зависит от длины полипептидной цепи. Белки дают сине-фиолетовое окрашивание; продукты их гидролиза (поли- и олигопептиды) – красную или розовую окраску. Биуретовую реакцию дают не только белки, пептиды и полипептиды но и биурет (NH 2 -CO-NH-CO-NH 2) , оксамид (NH 2 -CO-CO-NH 2), гистидин.
Образующееся в щелочной среде комплексное соединение меди (II) с пептидными группами имеет следующее строение:


Нингидриновая реакция . В этой реакции растворы белка, полипептидов, пептидов и свободных α-аминокислот при нагревании с нингидрином дают синее, сине-фиолетовое или розово-фиолетовое окрашивание. Окраска в этой реакции развивается за счет α-аминогруппы.


Очень легко реагируют с нингидрином -аминокислоты. Наряду с ними сине-фиолетовый Руэмана образуют также белки, пептиды, первичные амины, аммиак и некоторые другие соединения. Вторичные амины, например пролин и оксипролин, дают желтую окраску.
Нингидриновую реакцию широко используют для обнаружения и количественного определения аминокислот.
Ксантопротеиновая реакция. Эта реакция указывает на наличие в белках остатков ароматических аминокислот – тирозина, фенилаланина, триптофана. Основана на нитровании бензольного кольца радикалов этих аминокислот с образованием нитросоединений, окрашенных в желтый цвет (греческое «Ксантос» – желтый). На примере тирозина эту реакцию можно описать в виде следующих уравнений.
В щелочной среде нитропроизводные аминокислот образуют соли хиноидной структуры, окрашенные в оранжевый цвет. Ксантопротеиновую реакцию дают бензол и его гомологи, фенол и другие ароматические соединения.

Реакции на аминокислоты, содержащие тиоловую группу в восстановленном или окисленном состоянии (цистеин, цистин).
Реакция Фоля. При кипячении со щелочью от цистеина легко отщепляется сера в виде сероводорода, который в щелочной среде образует сульфид натрия:
В связи с этим реакции определения тиолсодержащих аминокислот в растворе подразделяют на два этапа:
Переход серы из органического состояния в неорганическое
Обнаружение серы в растворе
Для выявления сульфида натрия используют ацетат свинца, который при взаимодействии с гидроксидом натрия превращается в его плюмбит:
Pb(CH 3 COO) 2 + 2NaOH Pb(ONa) 2 + 2CH 3 COOH
В результате взаимодействия ионов серы и свинца образуется сульфид свинца черного или бурого цвета:
Na 2 S + Pb (ONa ) 2 + 2 H 2 O PbS (черный осадок) + 4 NaOH
Для определения серусодержащих аминокислот к исследуемому раствору добавляют равный объем гидроксида натрия и несколько капель раствора ацетата свинца. При интенсивном кипячении в течение 3-5 минут жидкость окрашивается в черный цвет.
Наличие цистина может быть определено с помощью этой реакции, так как цистин легко восстанавливается в цистеин.
Реакция Миллона:
Это реакция на аминокислоту тирозин.
Свободные фенольные гидроксилы молекул тирозина при взаимодействии с солями дают соединения ртутной соли нитропроизводного тирозина, окрашенной в розовато-красный цвет:

Реакция Паули на гистидин и тирозин . Реакция Паули позволяет обнаружить в белке аминокислоты гистидин и тирозин, которые образуют с диазобензолсульфоновой кислотой комплексные соединения вишнево-красного цвета. Диазобензолсульфоновая кислота образуется в реакции диазотирования при взаимодействии сульфаниловой кислоты с нитритом натрия в кислой среде:

К исследуемому раствору прибавляют равный объем кислого раствора сульфаниловой кислоты (приготовленного с использованием соляной кислоты) и двойной объем раствора нитрита натрия, тщательно перемешивают и сразу прибавляют соду (карбонат натрия). После перемешивания смесь окрашивается в вишнево-красный цвет при условии наличия гистидина или тирозина в исследуемом растворе.


Реакция Адамкевича-Гопкинса-Коля (Шульца - Распайля) на триптофан (реакция на индоловую группу). Триптофан реагирует в кислой среде с альдегидами, образуя окрашенные продукты конденсации. Реакция протекает за счет взаимодействия индольного кольца триптофана с альдегидом. Известно, что из глиоксиловой кислоты в присутствии серной кислоты образуется формальдегид:
Р астворы,
содержащие триптофан, в присутствии
глиоксиловой и серной кислот дают
красно-фиолетовое окрашивание.
астворы,
содержащие триптофан, в присутствии
глиоксиловой и серной кислот дают
красно-фиолетовое окрашивание.
Глиоксиловая кислота всегда присутствует в небольшом количестве в ледяной уксусной кислоте. Поэтому реакцию можно проводить, используя уксусную кислоту. При этом к исследуемому раствору добавляют равный объем ледяной (концентрированной) уксусной кислоты и осторожно нагревают до растворения осадка.После охлаждения к смеси осторожно по стенке (во избежание смешивания жидкостей) добавляют объем концентрированной серной кислоты, равный добавленному объему глиоксиловой кислоты. Через 5-10 минут на границе раздела двух слоев наблюдают образование красно-фиолетового кольца. Если перемешать слои, содержимое посуды равномерно окрасится в фиолетовый цвет.
К
 онденсация
триптофана с формальдегидом:
онденсация
триптофана с формальдегидом:
Продукт конденсации окисляется до бис-2-триптофанилкарбинола, который в присутствии минеральных кислот образует соли, окрашенные в сине-фиолетовый цвет:


7) Классификация белков. Способы исследования аминокислотного состава.
Строгой номенклатуры и классификации белков до сих пор не существует. Названия белков дают по случайным признакам, чаще всего принимая во внимание источник выделения белка или же учитывая растворимость его в тех или иных растворителях, форму молекулы и др.
Классификация белков проводится по составу, по форме частиц, по растворимости, по аминокислотному составу, по происхождению и т.д.
1. По составу белки делят на две большие группы: простые и сложные белки.
К простым (протеинам) относят белки, дающие при гидролизе только аминокислоты (протеиноиды, протамины, гистоны, проламины, глутелины, глобулины, альбумины). В качестве примеров простых белков могут служить фиброин шелка, яичный сывороточный альбумин, пепсин и др.
К сложным (к протеидам) относят белки, составленные из простого белка и добавочной (простетической) группы небелковой природы. Группу сложных белков делят на несколько подгрупп в зависимости от характера небелкового компонента:
Металлопротеиды, содержащие в своем составе металлы (Fe, Си, Mg и др.), связанные непосредственно с полипептидной цепью;
Фосфопротеиды - содержат остатки фосфорной кислоты, которые сложноэфирными связями присоединены к молекуле белка по месту гидроксильных групп серина, треонина;
Гликопротеиды - их простетическими группами являются углеводы;
Хромопротеиды - состоят из простого белка и связанного с ним окрашенного небелкового соединения, все хромопротеиды биологически очень активны; в качестве простетических групп в них могут быть производные порфирина, изоаллоксазина и каротина;
Липопротеиды - простетическая группа липиды - триглицериды (жиры) и фосфатиды;
Нуклеопротеиды - белки, состоящие из простого белка и соединенной с ним нуклеиновой кислоты. Эти белки играют колоссальную роль в жизнедеятельности организма и будут рассмотрены ниже. Они входят в состав любой клетки, некоторые нуклеопротеиды существуют в природе в виде особых частиц, обладающих патогенной активностью (вирусы).
2. По форме частиц - белки делят на фибриллярные (нитеподобные) и глобулярные (сферические) (см. стр 30).
3. По растворимости и особенностям аминокислотного состава выделяют следующие группы простых белков:
Протеиноиды - белки опорных тканей (костей, хрящей, связок, сухожилий, волос, ногтей, кожи и т.д.). Это в основном фибриллярные белки с большой молекулярной массой (> 150000 Да), нерастворимые в обычных растворителях: воде, солевых и водно-спиртовых смесях. Они растворяются только в специфических растворителях;
Протамины (простейшие белки) - белки, растворимые в воде и содержащие 80-90% аргинина и ограниченный набор (6-8) других аминокислот, представлены в молоках различных рыб. Вследствие высокого содержания аргинина имеют основные свойства, их молекулярная масса сравнительно мала и примерно равна 4000-12000 Да. Они являются белковым компонентом в составе нуклеопротеидов;
Гистоны - хорошо растворимы в воде и разбавленных растворах кислот (0,1Н), отличаются высоким содержанием аминокислот: аргинина, лизина и гистидина (не менее 30%) и поэтому обладают основными свойствами. Эти белки в значительных количествах содержатся в ядрах клеток в составе нуклеопротеидов и играют важную роль в регуляции обмена нуклеиновых кислот. Молекулярная масса гистонов невелика и равна 11000-24000 Да;
Глобулины - белки, нерастворимые в воде и солевых растворах с концентрацией соли более 7%. Глобулины полностью осаждаются при 50%-ном насыщении раствора сульфатом аммония. Эти белки отличаются высоким содержанием глицина (3,5%), их молекулярная масса > 100000 Да. Глобулины - слабокислые или нейтральные белки (р1=6-7,3);
Альбумины - белки, хорошо растворимые в воде и крепких солевых растворах, причем концентрация соли (NH 4) 2 S0 4 не должна превышать 50 % от насыщения. При более высокой концентрации альбумины высаливаются. По сравнению с глобулинами эти белки содержат глицина в три раза меньше и имеют молекулярную массу, равную 40000-70000 Да. Альбумины имеют избыточный отрицательный заряд и кислые свойства (pl=4,7) из-за большого содержания глутаминовой кислоты;
Проламины - группа растительных белков, содержащаяся в клейковине злаковых растений. Они растворимы только в 60-80%-ном водном растворе этилового спирта. Проламины имеют характерный аминокислотный состав: в них много (20-50%) глутаминовой кислоты и пролина (10-15%), в связи с чем они и получили свое название. Их молекулярная масса более 100000 Да;
Глютелины - растительные белки нерастворимые в воде, растворах солей и этаноле, но растворимы в разбавленных (0,1Н) растворах щелочей и кислот. По аминокислотному составу и молекулярной массе сходны с проламинами, но аргинина содержат больше, а пролина меньше.
Способы исследования аминокислотного состава
Под действием ферментов пищеварительных соков белки расщепляются на аминокислоты. Были сделаны два важных вывода: 1) в состав белков входят аминокислоты; 2) методами гидролиза может быть изучен химический, в частности амнокислотный, состав белков.
Для изучения аминокислотного состава белков пользуются сочетанием кислотного (НСl), щелочного [Ва(ОН) 2 ] и, реже, ферментативного гидролиза или одним из них. Установлено, что при гидролизе чистого белка, не содержащего примесей, освобождаются 20 различных α-аминокислот. Все другие открытые в тканях животных, растений и микроорганизмов аминокислоты (более 300) существуют в природе в свободном состоянии либо в виде коротких пептидов или комплексов с другими органическими веществами.
Первый этап в определении первичной структуры белков заключается в качественной и количественной оценке аминокислотного состава данного индивидуального белка. Необходимо помнить, что для исследования нужно иметь определённое количество чистого белка, без примесей других белков или пептидов.
Кислотный гидролиз белка
Для определения аминокислотного состава необходимо провести разрушение всех пептидных связей в белке. Анализируемый белок гидролизуют в 6 мол/л НС1 при температуре около 110 °С в течение 24 ч. В результате такой обработки разрушаются пептидные связи в белке, а в гидролизате присутствуют только свободные аминокислоты. Кроме того, глутамин и аспарагин гидролизуются до глутаминовой и аспарагиновой кислот (т.е. разрывается амидная связь в радикале и от них отщепляется аминогруппа).
Разделение аминокислот с помощью ионообменной хроматографии
Смесь аминокислот, полученных кислотным гидролизом белков, разделяют в колонке с катионообменной смолой. Такая синтетическая смола содержит прочно связанные с ней отрицательно заряженные группы (например, остатки сульфоновой кислоты -SO 3 -), к которым присоединены ионы Na + (рис. 1-4).
В катионообменник вносят смесь аминокислот в кислой среде (рН 3,0), где аминокислоты в основном представляют катионы, т.е. несут положительный заряд. Положительно заряженные аминокислоты присоединяются к отрицательно заряженным частицам смолы. Чем больше суммарный заряд аминокислоты, тем прочнее её связь со смолой. Так, аминокислоты лизин, аргинин и гистидин наиболее прочно связываются с катионообменником, а аспарагиновая и глутаминовая кислоты - наиболее слабо.
Высвобождение аминокислот из колонки осуществляют вымыванием (элюированием) их буферным раствором с увеличивающейся ионной силой (т.е. с увеличением концентрации NaCl) и рН. При увеличении рН аминокислоты теряют протон, в результате уменьшается их положительный заряд, а следовательно и прочность связи с отрицательно заряженными частицами смолы.
Каждая аминокислота выходит из колонки при определённом значении рН и ионной силы. Собирая с нижнего конца колонки раствор (элюат) в виде небольших порций, можно получить фракции, содержащие отдельные аминокислоты.
(подробнее «гидролиз» см вопрос №10)
8) Химические связи в структуре белка.



9) Понятие об иерархии и структурной организации белков. (см. вопрос №12)
10) Гидролиз белка. Химизм реакции (ступенчатость, катализаторы, реагенты, условия протекания реакции) – полное описание гидролиза.
11) Химические превращения белков.
Денатурация и ренатурация
При нагревании растворов белков до 60-80% или при действии реагентов, разрушающих нековалентные связи в белках, происходит разрушение третичной (четвертичной) и вторичной структуры белковой молекулы, она принимает в большей или меньшей степени форму беспорядочного случайного клубка. Этот процесс называют денатурацией. В качестве денатурирующих реагентов могут быть кислоты, щелочи, спирты, фенолы, мочевина, гуанидинхлорид и др. Сущность их действия в том, что они образуют водородные связи с =NH и =СО - группами пептидного остова и с кислотными группами радикалов аминокислот, подменяя собственные внутримолекулярные водородные связи в белке вследствие чего вторичная и третичная структуры изменяются. При денатурации падает растворимость белка, он "свертывается" (например, при варке куриного яйца), утрачивается биологическая активность белка. На этом основано, например, применение водного раствора карболовой кислоты (фенола) в качестве антисептика. В определенных условиях при медленном охлаждении раствора денатурированного белка происходит ренатурация - восстановление исходной (нативной) конформации. Это подтверждает тот факт, что характер укладки пептидной цепи определяется первичной структурой.
Процесс денатурации отдельной белковой молекулы, приводящий к распаду её «жёсткой» трёхмерной структуры, иногда называют плавлением молекулы. Практически любое заметное изменение внешних условий, например, нагревание или существенное изменение pH приводит к последовательному нарушению четвертичной, третичной и вторичной структур белка. Обычно денатурация вызывается повышением температуры, действием сильных кислот и щелочей, солей тяжелых металлов, некоторых растворителей (спирт), радиации и др.
Денатурация часто приводит к тому, что в коллоидном растворе белковых молекул происходит процесс агрегации частиц белка в более крупные. Визуально это выглядит, например, как образование «белка» при жарке яиц.
Ренатурация - процесс, обратный денатурации, при котором белки возвращают свою природную структуру. Нужно отметить, что не все белки способны ренатурировать; у большинства белков денатурация необратима. Если при денатурации белка физико-химические изменения связаны с переходом полипептидной цепи из плотно упакованного (упорядоченного) состояния в беспорядочное, то при ренатурации проявляется способность белков к самоорганизации, путь которой предопределён последовательностью аминокислот в полипептидной цепи, то есть её первичной структурой, детерминированной наследственной информацией. В живых клетках данная информация, вероятно, является решающей для преобразования неупорядоченной полипептидной цепи во время или после её биосинтеза на рибосоме в структуру нативной молекулы белка. При нагревании двухцепочечных молекул ДНК до температуры около 100°C водородные связи между основаниями разрываются, и комплементарные цепи расходятся - ДНК денатурирует. Однако при медленном охлаждении комплементарные цепи могут вновь соединяться в регулярную двойную спираль. Эта способность ДНК к ренатурации используется для получения искусственных гибридных молекул ДНК.
Природные белковые тела наделены определенной, строго заданной пространственной конфигурацией и обладают рядом характерных физико-химических и биологических свойств при физиологических значениях температуры и рН среды. Под влиянием различных физических и химических факторов белки подвергаются свертыванию и выпадают в осадок, теряя нативные свойства. Таким образом, под денатурацией следует понимать нарушение общего плана уникальной структуры нативной молекулы белка, преимущественно ее третичной структуры, приводящее к потере характерных для нее свойств (растворимость, электрофоретическая подвижность, биологическая активность и т.д.). Большинство белков денатурирует при нагревании их растворов выше 50–60°С.
Внешние проявления денатурации сводятся к потере растворимости, особенно в изоэлектрической точке, повышению вязкости белковых растворов, увеличению количества свободных функциональных SH-групп и изменению характера рассеивания рентгеновских лучей. Наиболее характерным признаком денатурации является резкое снижение или полная потеря белком его биологической активности (каталитической, антигенной или гормональной). При денатурации белка, вызванной 8М мочевиной или другим агентом, разрушаются в основном нековалентные связи (в частности, гидрофобные взаимодействия и водородные связи). Дисульфидные связи в присутствии восстанавливающего агента меркаптоэтанола разрываются, в то время как пептидные связи самого остова полипептидной цепи не затрагиваются. В этих условиях развертываются глобулы нативных белковых молекул и образуются случайные и беспорядочные структуры (рис.)

Денатурация белковой молекулы (схема).
а - исходное состояние; б - начинающееся обратимое нарушение молекулярной структуры; в - необратимое развертывание полипептидной цепи.

Денатурация и ренатурация рибонуклеазы (по Анфинсену).
а - развертывание (мочевина + меркаптоэтанол); б - повторное свертывание.

1. Гидролиз белков: H+
[− NH2─CH─ CO─NH─CH─CO − ]n +2nH2O → n NH2 − CH − COOH + n NH2 ─ CH ─ COOH
│ │ │ │
Аминокислота 1 аминокислота 2
2. Осаждение белков:
а) обратимое
Белок в растворе ↔ осадок белка. Происходит под действием растворов солей Na+, K+
б) необратимое (денатурация)
При денатурации под действием внешних факторов (температура; механическое воздействие – давление, растирание, встряхивание, ультразвук; действия химических агентов – кислот, щелочей и др.) происходит изменение вторичной, третичной и четвертичной структур белковой макромолекулы, т.е её нативной пространственной структуры. Первичная структура, а, следовательно, и химический состав белка не меняются.
При денатурации изменяются физические свойства белков: снижается растворимость, теряется биологическая активность. В тоже время увеличивается активность некоторых химических групп, облегчается воздействие на белки протеолитических ферментов, а, следовательно, он легче гидролизуется.
Например, альбумин - яичный белок - при температуре 60-70° осаждается из раствора (свертывается), теряя способность растворяться в воде.
Схема процесса денатурации белка (разрушение третичной и вторичной структур белковых молекул)
3. Горение белков
Белки горят с образованием азота, углекислого газа, воды, а также некоторых других веществ. Горение сопровождается характерным запахом жженых перьев
4. Цветные (качественные) реакции на белки:
а) ксантопротеиновая реакция (на остатки аминокислот, содержащих бензольные кольца):
Белок + HNO3 (конц.) → желтое окрашивание
б) биуретовая реакция (на пептидные связи):
Белок + CuSO4 (насыщ) + NaOH (конц) → ярко-фиолетовое окрашивание
в) цистеиновая реакция (на остатки аминокислот, содержащих серу):
Белок + NaOH + Pb(CH3COO)2 → Черное окрашивание
Белки являются основой всего живого на Земле и выполняют в организмах многообразные функции.
Высаливание белков
Высаливанием называется процесс выделения белков из водных растворов нейтральными растворами концентрированных солей щелочных и щелочноземельных металлов. При добавлении больших концентраций солей к раствору белка происходит дегидратация белковых частиц и снятие заряда, при этом белки выпадают в осадок. Степень выпадения белков в осадок зависит от ионной силы раствора осадителя, размера частиц белковой молекулы, величины ее заряда, гидрофильности. Разные белки осаждаются при различных концентрациях солей. Поэтому в осадках, полученных путем постепенного повышения концентрации солей, отдельные белки находятся в различных фракциях. Высаливание белков является обратимым процессом, и после удаления соли белок вновь приобретает природные свойства. Поэтому высаливанием пользуются в клинической практике при разделении белков сыворотки крови, а также при изолировании, очистке различных белков.
Добавляемые анионы и катионы разрушают гидратную белковую оболочку белков, являющуюся одним из факторов устойчивости белковых растворов. Чаще всего применяются растворы сульфатов Na и аммония. Многие белки отличаются по размеру гидратной оболочки и величине заряда. Для каждого белка есть своя зона высаливания. После удаления высаливающего агента белок сохраняет свою биологическую активность и физико-химические свойства. В клинической практике применяется метод высаливания для разделения глобулинов (при добавлении 50% раствора сульфата аммония (NH4)2SO4 выпадает осадок) и альбуминов (при добавлении 100% раствора сульфата аммония (NH4)2SO4 выпадает осадок).
На величину высаливания оказывают влияние:
1) природа и концентрация соли;
2) рН-среды;
3) температура.
Главную роль при этом играют валентности ионов.
12) Особенности организации первичной, вторичной, третичной структуры белка.
В настоящее время экспериментально доказано существование четырёх уровней структурной организации белковой молекулы: первичной, вторичной, третичной и четвертичной структуры.